
Original post is here.
I have been reading Steven Jay Gould's Wonderful Life: The Burgess Shale and the Nature of History. The work in entirety is a controversial illustration of a quiet yet revolutionary change in biology, which was later disputed by the researchers who made the discoveries. A section from page 80 is most relevant to my post, where Gould discusses how the romanticism of field work and the "technocratic myths about machine-based novelty" have very little to do with the new discovery of disparity of forms in the Burgess Shale.
The myth of field work, for example, proclaims that the great alteration in ideas arise from new, pristine discoveries. At the end of the trail, after weeks of blood, sweat, toil and tears, the intrepid scientist splits a rock from the most inaccessible place on the map, and cries Eureka! as he spies the fossil that will shake the world. Since the Burgess [ed.: taxonomic] revision was preceded by two full seasons of field work, in 1966 and 1967, most people assume that discoveries of this expedition prompted the reinterpretation. Well, [Harry] Whittington and company did find some wonderful specimens, and a few new species, but old [Charles] Walcott, a maniacal collector, had been there first, and had worked for five full seasons. He therefore got most of the goodies. The expeditions[...] did spur Whittington into action, but the greatest discoveries were made in museum drawers in Washington [at the Smithsonian]-- by restudying Walcott's well-trimmed specimens. The greatest bit of "field work", as we shall see, occurred in Washington during the spring of 1973, when Whittington's brilliant and eclectic student Simon Conway Morris made a systematic search through all the drawers of Walcott's specimens, consciously looking for oddities because he had grasped the germ of the key insight about Burgess disparity.For the most part, taxonomy does not employ any fancy new methods. In fossils, the methods are much the same as they were 100 years ago. In insects, this is similar; we have devised some new tools to find characters in molecules, but the old visual microscope and hand drawing methods are tried and true. Most universities with a physics department have had a scanning electron microscope since the 1970s, and many biology departments have their own.
...
The Burgess revision did require a definite set of highly specialized methods, but the tools of this particular technology do not extend beyond ordinary light microscopes, cameras, and dental drills. Walcott missed some crucial observations because he didn't use these methods--but he could have employed all Whittington's techniques, had he ever found the time to ponder, and to recognize their importance. Everything that Whittington did to see farther and better could have been done in Walcott's day.
Likewise, most taxonomic work, be it on fossils, insects, soil invertebrates, or ocean plankton, does not take place in the field. Most collecting is in bulk samples, which are later sorted through, and all the items of interest are taken out by the investigator. The rest of the sample (if not tossed) is stored for future research. For example, the Field Museum of Natural History in Chicago, IL, has an extensive alcohol-stored collection of insect bulk samples from around the world. Over decades these are picked through, and discoveries are made. In addition, all museums have orphaned specimens, or specimens the collector thought were interesting but didn't have enough time to investigate. Given the diversity of life in insects, it is not surprising that any single researcher, specializing in a particular group, will overlook new species or genera in his or her picking. Just like the Burgess Shale fossils, many future discoveries are already waiting on a museum shelf.
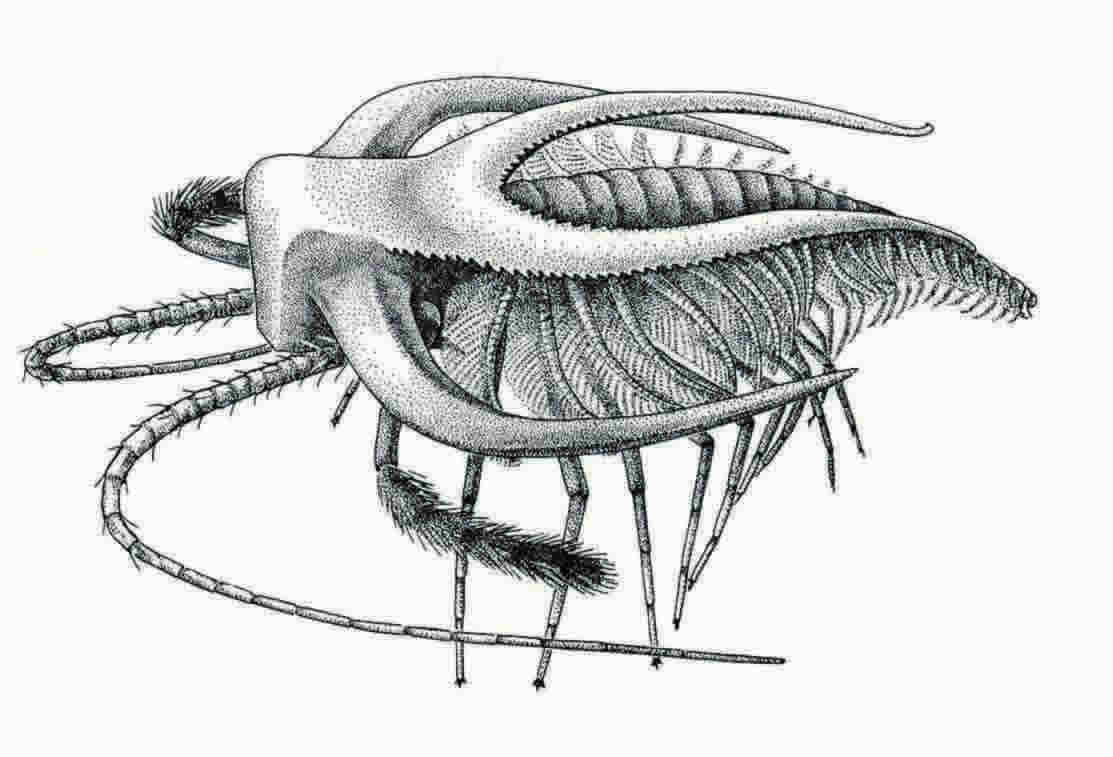
Drawing is a reconstruction of Marella, the most common Burgess Shale arthropod fossil. © Marianne Collins, from Gould's Wonderful Life (1989).


