
Imagine my surprise when I found this critter among the mix.
 The mystery fly, semi-left habitus. CHILE: PROVINCIA AYSEN: Municipalidad Cisnes, Parque Nacional Lago Copa, S. side of E. Lago Copa, unnamed 1st order stream, from cascade 200 m E. of Cliffs Lodge, S44.89155, W072.62070, 15-18.xii.2009.
The mystery fly, semi-left habitus. CHILE: PROVINCIA AYSEN: Municipalidad Cisnes, Parque Nacional Lago Copa, S. side of E. Lago Copa, unnamed 1st order stream, from cascade 200 m E. of Cliffs Lodge, S44.89155, W072.62070, 15-18.xii.2009.
If this was a fungus gnat, it was the strangest individual I had ever seen. First of all, the antennae are pectinate (feather-like), and there are few Diptera which have this character. This wouldn't eliminate Sciaroidea from the search, since there are genera in Ditomyiidae, Mycetophilidae, and Keroplatidae with species bearing pectinate antennae (Matile 1981). But then there are the other unusual characters.

The hindlegs are like nothing I had seen before. The tibiae are bowed and pressed forward in a grove against the femurs, so the tarsi lay close to the coxae. They resembled those of chalcid parasitoid wasps, which use those strong hind legs to capture prey for their young. The abdomen is short and conic. And the wings confuse the rest.

The wing veination looks as if it could have come from a fungus gnat, but the thick apical stigma and just how strong the veins are, how dark and heavy, reminds me more of a wasp wing. The halters mean it is obviously a true fly, but it looks like someone has glued on parts from other insect orders. I sent some pictures to Sciaroidea experts but the identity of this chimera continued to be a mystery.
I finally sent the above photos to Dalton de Souza Amorim, a South American fly expert, and he replied with a diagnosis he was nearly 100% sure of. But this was not a fungus gnat. It wasn't even in the Sciaroidea superfamily. This was a Canthyloscelidae.

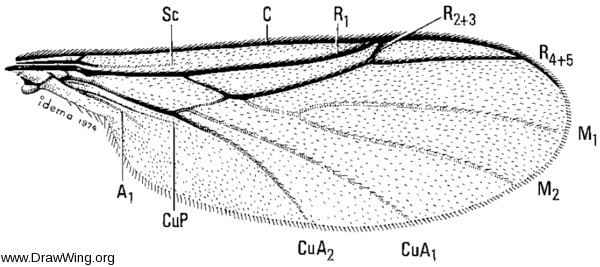
Wing of male Exiliscelis californiensis from the Synneuridae chapter of Manual of Nearctic Diptera. (Via DrawWing) If I didn't know better, I would say this came from a keroplatid.
I finally sent the above photos to Dalton de Souza Amorim, a South American fly expert, and he replied with a diagnosis he was nearly 100% sure of. But this was not a fungus gnat. It wasn't even in the Sciaroidea superfamily. This was a Canthyloscelidae.

Wing of male Exiliscelis californiensis from the Synneuridae chapter of Manual of Nearctic Diptera. (Via DrawWing) If I didn't know better, I would say this came from a keroplatid.
The family Canthyloscelidae is a small family of flies in the "lower" Diptera, with ~15 extant species in 4 genera, found native in all faunal regions except the East Palearctic and Afrotropics. There is disagreement as to whether it should be placed with the Bibiomorpha or Psychodomorpha; larval structures suggest the latter (Amorim 2008) while adult characters alone suggest the former. Canthyloscelidae used to be split into two or more families, the Synneuridae and Canthyloscelidae (sometimes previously combined under Hyperoscelidae (Hutson 1977)), but these were joined by the time of Haenii's canthyloscelid chapter in Contributions to a Manual of Palearctic Diptera (1997).
This particular species is Canthyloscelis pectinata, the only species in this family with the males bearing strongly pectinate antennae. It has been recorded from Argentina and Chile in very low numbers (Hutson 1977), which means this may be one of less than 20 specimens ever collected, a rare find (Amorim pers. com.).
 Exiscelis californiensis gathering on Potentilla flowers, Lowder Mountain, Oregon. (© 2011 Jake Hurlbert)
Exiscelis californiensis gathering on Potentilla flowers, Lowder Mountain, Oregon. (© 2011 Jake Hurlbert)
This particular species is Canthyloscelis pectinata, the only species in this family with the males bearing strongly pectinate antennae. It has been recorded from Argentina and Chile in very low numbers (Hutson 1977), which means this may be one of less than 20 specimens ever collected, a rare find (Amorim pers. com.).
Exiscelis californiensis gathering on Potentilla flowers, Lowder Mountain, Oregon. (© 2011 Jake Hurlbert)
In North America, there are two species of Canthyloscelidae in two genera, Synneuron decepiens and Exiscelis californiensis. S. decepiens is found at Northern latitudes across the continent, while E. californiensis is only found on the Northwest Coast. Both are found in "ancient forests" (Triplehorn and Johnson 2004). Very little is known about their biology; larvae live in decaying wood "permeated by [fungal] mycelia" (Peterson and Cook 1981), and this family's close relationship with Scatopsidae suggests they feed on the fungi and microorganisms associated with such habitats.
In all, it was a nice little treasure hunt. I think taking time to jump outside my comfort zone and identify something so strange to my experience is an excellent exercise. It improves my skill as a taxonomist and teaches me about groups beyond my research. But most of all, identifying something weird is /fun/. Finding something strange and new, even if it's just new to you, is one of the greatest pleasures of natural history research.
References
Haenni, J.-P. 1997. Family Canthyloscelidae. In Papp L. & Darvas B. (eds): Contributions to a Manual of Palaearctic Diptera. Nematocera and Lower Brachycera.. Vol. 2. Budapest: Science Herald. pp. 273–279.
Hutson, A. M. 1977. A revision of the families Synneuridae and Canthyloscelidae (Diptera). Bulletin of the British Museum (Natural History) Entomology 35(3):67-100.
Johnson, N. F., and C. A. Triplehorn. 2004. Borror and Delong's Introduction to the Study of Insects, 7th ed. Brooks Cole, St. Paul, MN.
Matile, L. 1981. A new Austrailian genus of Keroplatidae with pectinate antennae (Diptera: Mycetophiloidea). Journal of the Austrailian Entomological Society 20: 207-212.
Peterson, B. V., and E. F. Cook. 1981. Chapter 21. Synneuridae. Manual of Nearctic Diptera 1: 321-324.
In all, it was a nice little treasure hunt. I think taking time to jump outside my comfort zone and identify something so strange to my experience is an excellent exercise. It improves my skill as a taxonomist and teaches me about groups beyond my research. But most of all, identifying something weird is /fun/. Finding something strange and new, even if it's just new to you, is one of the greatest pleasures of natural history research.
References
Haenni, J.-P. 1997. Family Canthyloscelidae. In Papp L. & Darvas B. (eds): Contributions to a Manual of Palaearctic Diptera. Nematocera and Lower Brachycera.. Vol. 2. Budapest: Science Herald. pp. 273–279.
Hutson, A. M. 1977. A revision of the families Synneuridae and Canthyloscelidae (Diptera). Bulletin of the British Museum (Natural History) Entomology 35(3):67-100.
Johnson, N. F., and C. A. Triplehorn. 2004. Borror and Delong's Introduction to the Study of Insects, 7th ed. Brooks Cole, St. Paul, MN.
Matile, L. 1981. A new Austrailian genus of Keroplatidae with pectinate antennae (Diptera: Mycetophiloidea). Journal of the Austrailian Entomological Society 20: 207-212.
Peterson, B. V., and E. F. Cook. 1981. Chapter 21. Synneuridae. Manual of Nearctic Diptera 1: 321-324.


{kind=link}
{kind=link}